| CAS NO: | 301836-43-1 |
| 包装 | 价格(元) |
| 10mM (in 1mL DMSO) | 电议 |
| 10mg | 电议 |
| 50mg | 电议 |
| 200mg | 电议 |
| Physical Appearance | A solid |
| Storage | Store at -20°C |
| M.Wt | 398.41 |
| Cas No. | 301836-43-1 |
| Formula | C23H18N4O3 |
| Synonyms | Casein Kinase I Inhibitor;D 4476;D-4476 |
| Solubility | insoluble in H2O; ≥19.9 mg/mL in DMSO; ≥41.5 mg/mL in EtOH with gentle warming and ultrasonic |
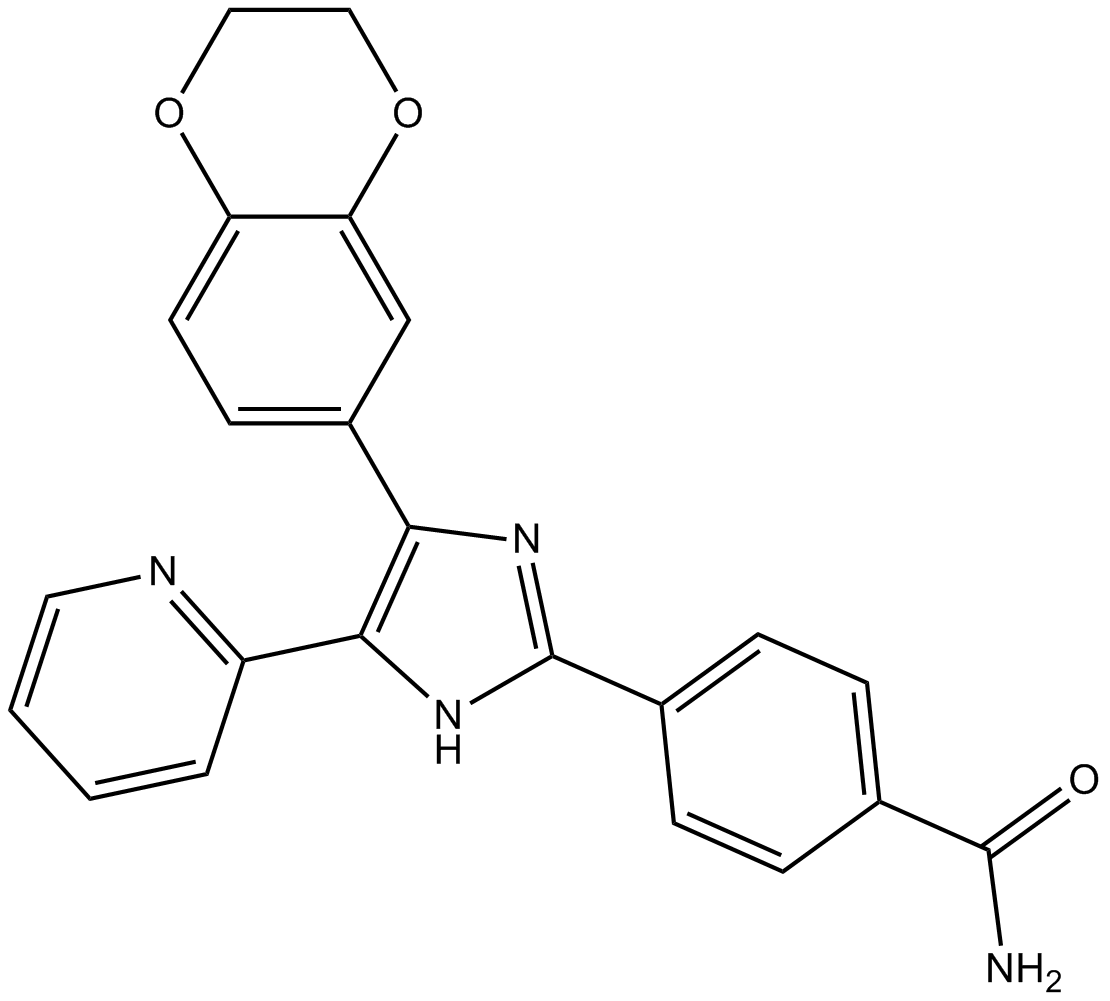
| Chemical Name | 4-[4-(2,3-dihydro-1,4-benzodioxin-6-yl)-5-pyridin-2-yl-1H-imidazol-2-yl]benzamide |
| Canonical SMILES | C1COC2=C(O1)C=CC(=C2)C3=C(NC(=N3)C4=CC=C(C=C4)C(=O)N)C5=CC=CC=N5 |
| 运输条件 | 蓝冰运输或根据您的需求运输。 |
| 一般建议 | 为了使其更好的溶解,请用37℃加热试管并在超声波水浴中震动片刻。不同厂家不同批次产品溶解度各有差异,仅做参考。若实验所需浓度过大至产品溶解极限,请添加助溶剂助溶或自行调整浓度。溶液形式一般不宜长期储存,请尽快用完。 |
D4476是一种相对特异性的和细胞穿透性的CK1和ALK5抑制剂,在体外0.1 mM ATP存在时的IC50值分别是0.3 和0.5 μM。[1]
CK1(酪蛋白激酶1)磷酸化多位点磷酸化结构域(MPDS)中靠近另一种磷酸化丝氨酸的丝氨酸残基。D4476可能是一种ATP竞争性的CK1抑制剂,其对SAPK2a/p38的抑制作用较弱并且对PKB或SGK无抑制作用。在H4IIE细胞中,FOXO1a的位点特异性磷酸化(Ser322和Ser325)分别被50和125 μM的D4476抑制,从而降低了核排斥的初始速率[1]。在Hela细胞中,CK1引起的RhoB磷酸化(Ser185)被50 μM的D4476抑制,然后RhoB依赖性肌动蛋白纤维形成随后被激活[2]。已观察到D4476治疗在HCT116细胞中产生部分的p53依赖性生长阻滞。[3]
参考文献:
1. G. Rena, J. Bain, M. Elliott and P. Cohen, EMBO Rep 2004, 5, 60-65.
2. V. Tillement, I. Lajoie-Mazenc, A. Casanova, C. Froment, M. Penary, D. Tovar, R. Marquez, B. Monsarrat, G. Favre and A. Pradines, Exp Cell Res 2008, 314, 2811-2821.
3. A. S. Huart, N. J. MacLaine, D. W. Meek and T. R. Hupp, J Biol Chem 2009, 284, 32384-32394.
| 细胞实验 [1,2]: | |
细胞系 | H4IIE、Hela、HCT116细胞系 |
溶解方法 | 在DMSO中的溶解度> 19.9 mg/mL。为了获得更高浓度,可以将离心管在37℃加热10分钟和/或在超声波浴中震荡一段时间。原液可以在-20℃以下储存几个月。 |
反应条件 | H4IIE细胞: 0-150 μM孵育1小时;Hela细胞:50 μM;HCT116细胞:5– 40 μM孵育48小时 |
应用 | 在H4IIE肝癌细胞中,D4476能够特异性抑制其MPD中内源性叉头盒转录因子O1a(FOXO1a)的Ser322和Ser325磷酸化,而不影响其他部位的磷酸化。 还发现,尽管HCT116细胞经D4476处理后会造成部分p53依赖性生长停滞,但D4476诱导的p53非依赖性凋亡表明MDM2-CK1复合物在维持E2F-1抗凋亡信号通路中的关键作用。 |
References: [1] Rena G, et al. D4476, a cell-permeant inhibitor of CK1, suppresses the site-specific phosphorylation and nuclear exclusion of FOXO1a. EMBO Rep. 2004 Jan;5(1):60-5. [2] Huart AS, MacLaine NJ, Meek DW, Hupp TR.CK1alpha plays a central role in mediating MDM2 control of p53 and E2F-1 protein stability. J Biol Chem. 2009 Nov 20;284(47):32384-94. | |
 m.cnreagent.com
m.cnreagent.com